Тепло как экологический фактор
Жизнедеятельность любого вида протекает в определенных интервалах температур. При этом прослеживается зона оптимума, минимума и максимума. В зоне минимума или максимума происходит затухание активной деятельности организма. В первом случае низкие температуры (холод), а во втором высокие (жара) приводят к нарушению жизненных процессов. За пределами этих крайних температур - летальная зона, в которой наступает необратимый процесс отмирания растения. Следовательно, температуры определяют границы жизни.
По причине неподвижного образа жизни высшие растения выработали большую выносливость к суточным и сезонным (годовым) колебаниям температур. Многие лесообразующие породы нашей тайги - сосна сибирская (сибирский кедр - Pinus sibirica), лиственница даурская (Larix dahurica) и др. - выдерживают понижения температуры до - 50°С и ниже и летнее тепло до +25°С и выше. Годовая амплитуда достигает 75°, а иногда 85 - 90°С. Виды растений, выдерживающие большие перепады температур, называют эвритермными (от греч. эври - широкий, терма - теплота) в отличие от стенотермных (стенос - узкий).
Дифференциация тепла на нашей планете - основа широтной зональности и высотной поясности растительности и почв.
Вследствие уменьшения от экватора к полюсам высоты солнцестояния и угла падения лучей изменяется количество тепла. Так, среднегодовая температура около экватора составляет +26,2°С, возле 30° с. ш. она уже равна +20,3°С, а при 60° с. ш. снижается до - 1°С.
Помимо среднегодовой температуры данной местности, важное значение в жизни организмов имеет наиболее высокая и наиболее низкая температура (абсолютный максимум и абсолютный минимум), наблюдаемая в данной климатической зоне, средняя температура самого теплого и самого холодного месяца. Так, продолжительность вегетационного периода в тундре (т. е. выше 70° с. ш.) составляет всего 1,5 - 2,5 мес при средней температуре 10 - 12°С. Растительный покров формируют кустарничковые жизненные формы ив (Salix arctica, S. herbacea, S. polaris) и берез (Betula папа, В. rotundifolia). Травянистые жизненные формы представляют мхи, лишайники, осоки и злаки. В условиях Крайнего Севера покрытосеменные почти полностью исчезают - здесь господствуют лишайники. В подзоне лесотундры (65 - 70° с. ш.) вегетационный период равен 2 - 3 мес, при средней температуре 12 - 14°С растительный покров намного богаче, в том числе по разнообразию видов древесных пород: сосен, елей. Тайга, иначе зона хвойных лесов, имеет вегетационный период 3 - 5 мес, средняя температура 14 - 16°С. В южной части зоны, где преобладают хвойно-широколиственные леса, вегетация длится 4 - 5 мес, средняя температура 15 - 16°С. В зоне широколиственных лесов (40 - 50° с. ш.) вегетационный период 5 - 6 мес, средняя температура 16 - 18°С. Здесь произрастают многие широколиственные деревья (дуб, бук, ясень, клены, липа и др.). Резким контрастом этим зонам выступает, например, зона дождевых тропических лесов (0 - 15° с. и ю. ш.). Вегетационный период здесь круглогодичный со средней температурой 25 - 28°С и часто не дифференцирован на сезоны. Исключительно важной особенностью тропических районов является ситуация, когда разница между средними температурами самого теплого и самого холодного месяца менее контрастна, чем суточные колебания.
Рост растений непосредственно связан с температурным фактором. Конечно, зависимость разных видов от температуры колеблется в широких пределах. Четко различаются термофильные (от греч. термо - теплота, филос - любовь) растения и их антиподы - холодовыносливые, или криофильные (от греч. криос - холод). А. Декандоль (1885) выделяет группы гекистотермных, микротермных, мезотермных и мегатермных растений (от греч. гекистос - холод, микрос - малый, мегас - большой). Процесс роста особей каждого вида имеет свои минимальные, оптимальные и максимальные температурные точки, поэтому в каждом случае зависимость роста от температуры легко выразить графически.
Не менее наглядно влияние температур на распространение отдельных видов растений и их группировок. Давно уже установлена взаимосвязь географического распространения отдельных видов с изотермами. Как известно, виноград созревает в пределах изотермы со средней температурой в течение 6 мес (апрель - сентябрь) 15°С. Распространение дуба черешчатого на север ограничено годовой изотермой 3°С; северная граница плодоношения фцниковой пальмы совпадает с годовой изотермой (18 - 19°С).
Однако в целом ряде случаев распространение растений обусловлено не только температурами. Так, изотерма 10°С проходит с запада на восток через Ирландию, ФРГ (Карлсруэ), Австрию (Вена), Украину (Одесса). Но названные местности имеют достаточно различный видовой состав природного растительного покрова и представляют возможность интродукции и возделывания разнообразного набора культур. Так, в Ирландии часто не вызревают зерновые культуры. В ФРГ, а тем более в той же Ирландии не вызревают многие тыквенные (арбузы, дыни), хотя в открытом грунте произрастают камелии и пальмы. В Карлсруэ в открытом грунте растут плющ и падуб, иногда вызревает и виноград. В районе Одессы возделывают дыни и арбузы, но плющ и камелии не выдерживают низких температур зимы. Таких примеров можно привести много. Средние температуры, тем более в отрыве от других факторов среды, не являются надежным показателем (индикатором) возможности интродукции и возделывания интересующей нас культуры. Суть дела в том, что разные виды растений характеризуются разной продолжительностью вегетационного периода. Поэтому в отношении температуры необходимо учитывать как продолжительность периода благоприятных температур для нормального развития растений, так и время наступления и продолжительность действия минимальных температур (то же в отношении максимальных).
В экологической и растениеводческой литературе для оценки тепловых ресурсов вегетационного периода широко используют сумму активных температур. Она служит хорошим показателем для оценки потребности растений в тепле и дает возможность определить район возделывания той или иной сельскохозяйственной культуры. Сумма активных температур состоит из суммы положительных среднесуточных температур за период, когда она выше 10°С. В районах, где сумма активных температур равна 1000 - 1400°С, можно возделывать ранние сорта картофеля, корнеплоды; там, где эта сумма достигает 1400 - 2200°С, - хлебные злаки, картофель, лен и др.; сумма активных температур 2200 - 3500°С соответствует зоне интенсивного плодоводства; при сумме температур более 4000°С успешно возделывают субтропические многолетники.
Организмы, жизнедеятельность которых и температура тела зависят от тепла, поступающего из окружающей среды, называют пойкилотермными (от греч. пойкилос - пестрый, разнообразный). К ним относят все растения, микроорганизмы, беспозвоночных животных и некоторые группы хордовых. Температура их тела зависит от среды. Вот почему экологическая роль тепла в жизни всех систематических групп растений и названных групп животных имеет первостепенное значение. Высокоорганизованных животных (птицы и млекопитающие) относят к группе гомойотермных (от греч. гомойос - равный), у которых температура тела постоянная, поскольку поддерживается за счет собственного тепла.
Известно, что протопласт клеток живых организмов способен нормально функционировать в интервале температур между 0 и 50°С. Только организмы, которые имеют специальные приспособления, могут выдерживать экстремальные температуры в течение длительного времени.
Физиологами установлены оптимальные и критические температуры дыхания и других функций. Оказывается, нижний предел температуры дыхания у зимующих органов (почки, хвоя) - минус 20 - 25°С. При повышении температуры интенсивность дыхания возрастает. Температуры свыше плюс 50°С разрушают белково-липидный комплекс поверхностного слоя цитоплазмы. Это приводит к потере клетками осмотических свойств.
В ряде районов СССР часто наблюдается массовая гибель растений от слишком низких температур. Катастрофическое их действие в наибольшей степени сказывается в малоснежные зимы преимущественно на озимых хлебах. Губительно также внезапное похолодание весной, когда растения трогаются в рост (поздневесенние заморозки). Нередко от холода гибнут не только интродуцированные вечнозеленые древесные, например цитрусовые, но и листопадные растения. Н. Максимов, изучая механизм действия низких температур, пришел к выводу, что причина гибели растений объясняется обезвоживанием цитоплазмы. В межклетниках ткани происходит кристаллизация воды. Кристаллы льда оттягивают воду из клеток. Они же механически повреждают органеллы клеток. Критический момент наступает именно с появлением кристаллов льда внутри клеток. В настоящее время выявляют уже целые природные группы морозоустойчивых растений. К ним можно отнести хвойные вечнозеленые деревья и кустарники, а также бруснику, вереск и др. Среди травянистых многолетников также выявлено немало морозоустойчивых растений, способных переживать суровую зиму.
Работами И. Туманова (1961) доказано, что в период зимнего покоя растения могут выдерживать очень низкие температуры. Так, побеги черной смородины (Ribes nigrum) при медленном снижении температур до минус 253°С (это температура, близкая к абсолютному нулю) могут сохранять свою жизнедеятельность.
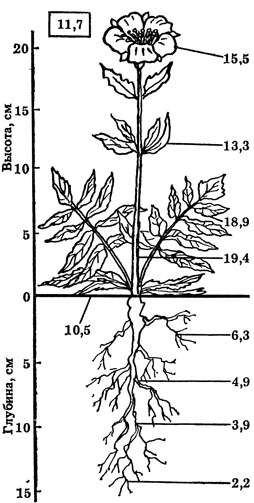
Рис. 18. Температура разных органов розеточного растения новосиверсии (Novosiversia gracilis) (по Б. Тихомирову), °С
Большинство видов растений характеризуется индивидуальными реакциями на температуры. Так, весной прорастание зерновок у ржи начинается при температуре плюс 1 - 2°С, у красного клевера - при 1°С, у люпина желтого - при 4 - 5°, у риса - при плюс 10 - 12°С. Соответственно оптимальные температуры для ржи составляют 25°С, для риса 30 - 32°С, для клевера 30°С, для люпина плюс 28°С.
Для нормального роста и развития растений необходима соответствующая температура для надземных и подземных органов. Например, лен нормально развивается при температуре корня примерно в 2 раза ниже (10°С), чем надземных органов (22°С). В ходе онтогенеза потребность растений в тепле заметно изменяется. Б. Тихомиров (1963) показал, насколько значительно изменяется температура органов тела растения в зависимости от их местонахождения (почва, воздух) и ориентировки по отношению к солнечным лучам (рис. 18). Экспериментально установлено, что прорастание семян рапса, сурепицы, пшеницы, овса, ячменя, клевера, люцерны и др. наблюдается при температуре 0 плюс 2°С, тогда как для появления всходов требуются более высокие температуры - 3 - 5°С. На многих видах континентальных растений благоприятно сказывается суточный термопериодизм, когда амплитуда ночных и дневных температур составляет от 5 - 10 до 15°С.
Опытные данные свидетельствуют также, что растениям умеренных широт для нормального онтогенетического развития необходимы осенние пониженные температуры - сезонный термопериод, а также суточный термопериод. Суть его заключается в том, что многие растения более успешно развиваются при пониженных ночных температурах. Например, томаты лучше развиваются, если дневная температура воздуха достигает 26°С, а ночная температура 17 - 18°С.
Мегатермные растения тропического происхождения, например сахарный тростник (Saccharum officinarum), нуждаются в высоких температурах в течение всей жизни. Следовательно, температурный фактор влияет на растения на всех этапах их роста и развития. Причем в разные периоды различные растения нуждаются в определенных температурных условиях. Для большинства однолетних растений, например ячменя, овса и др., прослеживается общая закономерность: на ранних фазах развития температура должна быть ниже, чем на более поздних.
Наибольшей выносливостью к сверхвысоким температурам отличаются растения жарких и сухих районов - эуксерофиты, а также многие суккуленты, например кактусы и очитки (см. следующий раздел). Это свойственно и растениям засоленных (особенно сульфидами и хлоридами) почв. Они, как показал еще X. Люденгорд (1925, 1937), сохраняют жизнеспособность даже при температуре 70°С. Хорошо переносят высокие температуры сильно обезвоженные семена и плоды. Именно на этом их свойстве основан, например, известный метод борьбы с возбудителем пыльной головни пшеницы (Ustilago tritici). При термообработке пораженных семян гриб, будучи етенотермным, погибает, тогда как зародыш зерновки остается жизнеспособным.
Труднее решить вопрос о влиянии температуры на изменение структуры самого растения, на его морфологию. Наблюдения в природе и экспериментальные дают различные объяснения. В самом деле, такое приспособление, как сильное опушение почечных чешуй и листьев является комплексным, оно служит защитой не только от яркого света, но и от высоких температур, а также от излишнего испарения влаги. Яркий блеск глянцевитых листьев, параллельное расположение листовой пластинки к солнечным лучам, войлочное опушение - все это, несомненно, предотвращает перегрев листа, а также излишнюю транспирацию.
Основатель экологии растений Е. Варминг (1895) наглядно продемонстрировал влияние температуры на формирование приземистых и розеточных форм растений в Арктике и в высокогорьях альпийского и субнивального пояса. Речь идет не только о травянистых бесстебельных, розеточных формах вроде девясила корнеглавого (Inula rhizocephala), но и о древесных жизненных формах - карликовой березе, можжевельнике туркестанском, кедровом стланике и др. Стелющиеся и подушечные формы растений (например, Minuartia arctica) наиболее приспособлены к условиям жизни у самой поверхности почвы под прикрытием снежного покрова. Когда же снега нет, в припочвенном слое воздуха на высоте до 15 - 20 см сохраняется наиболее высокая температура и сила ветра минимальна. К тому же внутри подушки создается особый микроклимат, и колебания температур здесь гораздо менее выражены, чем вне ее.
Таким образом, на формирование приземистых форм температурный фактор может действовать и непосредственно, и косвенно, т. е. путем нарушения водоснабжения и минерального питания. Помимо температур, определенное влияние (также прямое и косвенное) оказывают ветер и другие факторы.
Наиболее велика роль прямого влияния температур в процессе геофилизации растений. Под геофилизацией понимают погружение нижней (базальной) части растения в почву (сначала гипокотиля, затем эпикотиля, первого междоузлия и т. д.). Этот феномен свойствен преимущественно покрытосеменным растениям. Именно в ходе исторического развития покрытосеменных геофилизации играла видную роль в трансформации жизненных форм от деревьев к травам. С погружением в почву основания побегов интенсивно развивается система придаточных корней, корневищ, столонов и других органов вегетативного размножения. Геофилизация была необходимой предпосылкой появления разнообразных подземных органов растений, особенно органов вегетативного размножения. Это дало покрытосеменным большие преимущества в борьбе за существование, за господство на континентах Земли.
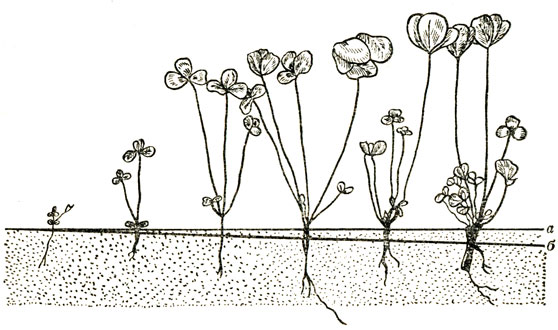
Рис. 19. Геофилизации (втягивание в почву) подсемядольного колена клевера лугового (Trifolium pratense) (по П. Лисицину): а - поверхность почвы; б - глубина втягивания
В онтогенезе многих покрытосеменных геофилизация растений осуществляется с помощью особых втягивающих (контрактильных) корней. Интересные экспериментальные исследования по геснфилизации провел профессор ТСХА П. Лисицин. Он выяснил, что втягивание в почву базальной части растения распространено гораздо шире, чем ранее предполагалось (рис. 19). Для озимых сельскохозяйственных культур геофилизация улучшает условия зимовки, для яровых, например гречихи, - условия водоснабжения.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://geoman.ru/ 'Физическая география'