Свет как экологический фактор
Жизнь нашей планеты во всем ее разнообразии обусловливается энергией солнечной радиации. Солнечная энергия приводит в движение сложный механизм воздушных и морских течений. Под ее воздействием происходит испарение воды и свершается непрерывный влагооборот. Велика роль света в жизни растений. Только очень немногие виды осуществляют синтез не за счет света, а за счет химической энергии неорганических соединений. Таковы серобактерии, железобактерии, нитрифицирующие бактерии. Создание органического вещества хлорофиллоносными растениями осуществляется за счет лучистой энергии солнца. Однако свет необходим и гетеротрофным растениям, например грибам-сапрофитам и даже паразитам.
Возделываемые культуры, которые особо требовательны к свету, например рис (Oryza sativa), при его недостатке явно задерживает свое развитие. Формирование высокопроизводительных древостоев многих лесообразующих пород и плодовых насаждений также находится в большой зависимости от интенсивности солнечной энергии. Сахаристость свеклы (Beta vulgaris) прямо зависит от интенсивности лучистой энергии солнца в течение вегетационного периода. Известно, что лен (Linum usitatissimum) и конопля (Cannabis sativa) в условиях короткого светового дня синтезируют значительное количество масла, а в условиях длинного убыстряют формирование лубяных волокон. Реакция растений на длину дня и ночи проявляется в ускорении или задержке развития. Следовательно, действие света на растение избирательно и неоднозначно. Значение освещенности как экологического фактора для организма определяется продолжительностью, интенсивностью и длиной волн светового потока.
На границе земной атмосферы с космосом радиация составляет от 1,98 до 2 кал/см2 в минуту. Эту величину называют солнечной постоянной. К поверхности Земли при разных погодных условиях доходит только 42 - 70% солнечной постоянной. Солнечная радиация, проходя через атмосферу - "окно прозрачности", претерпевает целый ряд изменений. Поэтому она существенно изменена не только в количественном отношении, но и по составу. Коротковолновая радиация поглощается озоновым экраном, который расположен на высоте около 25 км, и кислородом воздуха. Инфракрасные лучи поглощаются в атмосфере водяными парами и углекислым газом. В результате этого происходит нагревание воздуха. Остальная часть лучистой энергии достигает поверхности Земли в виде прямой или рассеянной радиации (рис. 15). Количество достигающей поверхности Земли радиации обусловлено географической широтой местности, продолжительностью дня, прозрачностью атмосферы и углом падения солнечных лучей. В ясные солнечные дни лучистая энергия, достигающая поверхности Земли, состоит на 45% из видимого света (380 - 720 нм) и на 45% из инфракрасного излучения, только 10% приходится на ультрафиолетовое излучение. Совокупность прямой и рассеянной солнечной радиации составляет суммарную радиацию.
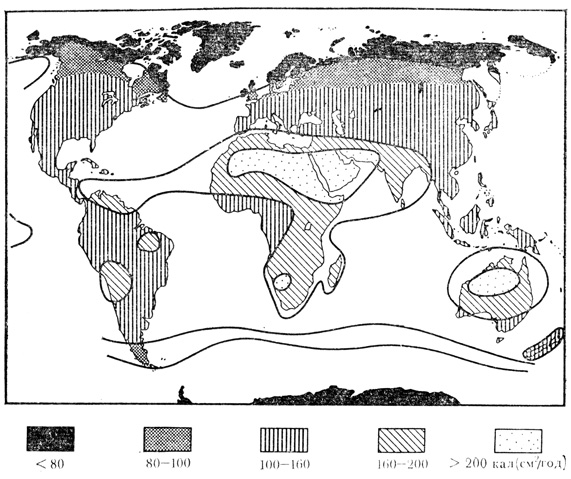
Рис. 15. Солнечная радиация, падающая на поверхность Земли (по В. Лархеру)
В ясные дни рассеянная радиация составляет от 1/3 до 1/8 суммарной радиации, тогда как в облачные дни рассеянная радиация составляет 100%. Значительное влияние на радиационный режим оказывает запыленность атмосферы. Так, в некоторых городах из-за загрязнения атмосферы освещенность может снижаться до 15% и более, чем за городом.
Освещенность на поверхности Земли варьируется в широких пределах. Все зависит от высоты стояния солнца над горизонтом, т. е. от угла падения солнечных лучей, длины дня и условий погоды, от прозрачности атмосферы. Интенсивность света также колеблется в зависимости от времен года и времени суток. Неравноценно в отдельных районах Земли и качество света, например соотношение длинноволновых (красных) и коротковолновых (синих и ультрафиолетовых) лучей. Как известно, коротковолновые лучи больше, чем длинноволновые, поглощаются и рассеиваются атмосферой. Поэтому в горных местностях всегда больше коротковолновой солнечной радиации.
Установлено, что в высоких широтах преобладает рассеянная радиация, а под тропиками - прямая. Рассеянная радиация содержит в полдень желто-красных лучей до 60%, прямая же - 30 - 40%. Поскольку фотосинтетически активная радиация (ФАР) представлена участком спектра между длиной волны 380 и 710 нм и максимальна в области оранжево-красных лучей (600 - 680 нм), то естественно, что коэффициент использования растениями рассеянной радиации выше. Отметим, что свет даже в высоких северных широтах за счет увеличения длины дня не ограничивает жизнедеятельности растений. Л. Иванов рассчитал, что даже на о-ве Шпицберген солнечной радиации достаточно (порядка 20 тыс. килоджоулей или 5 тыс. ккал на 1 га), чтобы получить необходимый урожай сухой растительной массы.
У разных видов растений и разных растительных группировок потребность к свету различна. Фундаментальные экспериментальные исследования по установлению коэффициента использования солнечного света древесными растениями проводил Л. Иванов. Известный московский растениевод А. Дояренко установил, что большинство сельскохозяйственных травянистых растений имеет коэффициент использования света для фотосинтеза 2 - 2,5%. Он приводит конкретные величины: свекла кормовая - 1,91%, вика - 1,98, клевер - 2,18, картофель - 2,48, рожь - 2,42, пшеница - 2,68, овес - 2,74, лен - 3,61, люпин - 4,79%. Установлено также, что разные виды растений неодинаково реагируют на интенсивность освещения. Таким образом очевидно, что разным растениям для нормальной вегетации необходимо различное световое довольствие (L), т. е. доля в процентах от полной ФАР. Это позволяет выделить в отношении потребности к свету три экологические группы растений: 1) световые растения, или гелиофиты (от греч. гелиос - солнце) (Lопт=100%, Lмин=70%), - растения открытых пространств. Сюда относятся, например, ковыли, большинство культивируемых растений: сахарная свекла, картофель и др.; 2) теневыносливые растения, или гемисциофиты. Они могут расти при L=100%, но могут переносить и большое затенение: ежа сборная (Dactylis glomerata), например, способна вегетировать в диапазоне L от 100 до 2,5%; 3) тенелюбивые растения - сциофиты (от греч. скиа - тень) не переносят полного освещения, их Lмакс всегда меньше 100%: кислица (Oxalis acetosella), седьмичник (Trientalis europea) и др. Ввиду особой структуры листья сциофитов при слабой интенсивности света способны ассимилировать углекислый газ не менее эффективно, чем листья гелиофитов.
Из растительных сообществ лесные наиболее активно трансформируют состав солнечного света, и до поверхности почвы доходит очень малая часть первоначальной солнечной радиации. Известно, что листовая поверхность древостоя поглощает около 80% падающей ФАР, еще 10% отражается и только 10% проникает под полог леса. Следовательно, полная и проникшая сквозь полог древесных растений радиация имеет не только количественное, но и существенные качественные различия. Растения, обитающие под пологом других, довольствуются лишь долей полного освещения. Это сциофиты и гемисциофиты. Так, если у тенелюбивой кислицы (Oxalis acetosella) максимум интенсивности фотосинтеза достигается при 1/10 полного дневного освещения, то у светолюбивых видов максимум наступает примерно при 1/2. К тому же светолюбивые менее приспособлены к существованию при слабом освещении по сравнению с тенелюбивыми и теневыносливыми. Нижний предел, при котором могут расти лесные зеленые мхи, - 1/90 полного дневного освещения. В дождевых тропических лесах встречаются еще более сциофильные виды, вегетирующие при 1/120 полного освещения. Удивительны в этом отношении некоторые мхи: шистостега перистая (Schistostega peunata и др.) - это растения темных пещер, вегетирующие при V2000 полного освещения.
Адаптация растений к световому режиму. Каждая географическая местность характеризуется определенным световым режимом. Важнейшими элементами светового режима, определяющими направление адаптации растений, являются интенсивность радиации, спектральный состав света, продолжительность освещения (длина дня и ночи).
Фотопериодизм. Продолжительность солнечного дня постоянна лишь на экваторе. Здесь день, как и ночь, длится 12 ч. Длительность солнечного дня в течение летнего периода возрастает от экватора в направлении обоих полюсов; на полюсе, как известно, целое лето длится полярный день, а зимой - полярная ночь. Реакцию растения на сезонные изменения длины дня и ночи называют фотопериодизмом.
В процессе растениеводческой практики давно обратили внимание, что сельскохозяйственные растения разного происхождения неодинаково реагируют на длину солнечного дня. Одни виды обособились как растения длинного дня, другие - короткого, третьи на длину дня заметно не реагируют. Общеизвестно, что в условиях длинного дня формируется высокий урожай пшеницы, ржи, овса и ряда кормовых злаков; к растениям длинного дня относятся также картофель, цитрусовые и ряд других овощных и плодовых культур. Продолжительное освещение этих растений вызывает более быстрое прохождение фаз развития плодов и семян.
С другой стороны, известны растения короткого дня, например просо (Panicum miliaceum), сорго (Sorghum cernuum), рис и др. Скорость прохождения этапов развития растений короткого дня при длительном освещении замедляется. Стимуляция быстрейшего цветения и плодоношения достигается укорачиванием времени освещения.
Таким образом, для практики земледелия при интродукции сельскохозяйственных растений очень существен учет продолжительности дня и ночи. Растения низких широт (южные растения) часто бывают растениями короткого дня. При интродукции в высокие широты, т. е. в условиях длинного дня, они развиваются медленно, зачастую не вызревают, а иногда даже не зацветают, как, например, конопля (Cannabis sativa). К этой же группе можно отнести топинамбур (Helianthus tuberosus), выращиваемый в Московской обл. Значит, продолжительность дня и ночи может определять границы распространения и возможной интродукции отдельных видов: "южных" на север, "северных" на юг. К числу нейтральных в отношении длины дня относятся томаты и др.
В ходе изучения фотопериодизма и фотохимических реакций выяснено, что у растений длинного дня в весенне-летний период (когда природе этих растений соответствует длинный солнечный день) явно убыстряется рост. Однако во второй половине лета, когда солнечный день сокращается, ростовые процессы явно замедляются. В результате у длиннодневных растений до наступления морозов не всегда успевает сформироваться комплекс покровных тканей - перидерма. Значит, длиннодневные многолетние культуры, возделываемые в условиях высоких широт, могут утратить зимостойкость. Это нужно иметь в виду при подборе ассортимента растений для возделывания в высоких широтах. Предпочтительнее в условиях длинного дня вводить однолетние культуры, не требующие перезимовки. Продвижению к северу некоторых культур, например клеверов, препятствуют не зимние морозы, а характер фотопериодических реакций. Именно так можно объяснить парадоксальный факт, что морозостойкость клеверов и люцерны более высока в средней полосе европейской части СССР, чем в северной.
Свет оказывает формирующее действие на растения. Это проявляется в размерах, форме и структуре (макро- и микроскопической) световых и теневых листьев (рис. 16), а также во влиянии на ростовые процессы. Важно отметить, что зависимость структуры листа (побега) от света не всегда прямая; листья (побеги), развивающиеся весной, формируются в соответствии с освещением не текущего года, а прошлого, т. е. когда закладывались почки. В этой связи И. Серебряков (1962) считал, что световая структура листа детерминирована уже в почке. Листья сохраняют эту структуру достаточно устойчиво даже при перенесении световых побегов в затенение. Большая высота, колоннообразная форма стволов, высокое расположение крон (очищение от сухих ветвей) характеризуют светолюбивые растения.
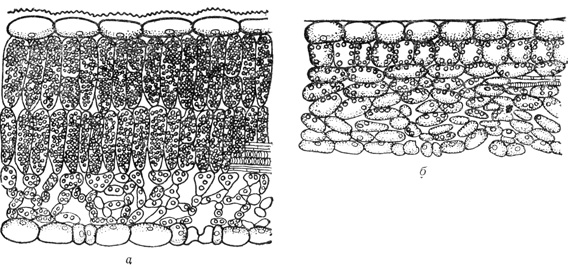
Рис. 16. Поперечные разрезы листьев сирени (Syringa): а - световой лист; б - теневой лист
Одной из реакций светолюбивых растений является затормаживание роста надземных побегов. Это приводит в одних случаях к сильной ветвистости, в других к розеточности. Отличает их и ряд других изменений структуры тела: мелколистность, увеличение мощности наружной стенки эпидермы и ее выростов (трихом и эмергениев), кутикулярного слоя и т. д.
Листовая мозаика и компасные растения. Одним из примеров приспособлений к свету является ориентировка листовой пластинки по отношению к солнечным лучам. Различают три способа ориентировки: 1) листовая пластинка ориентирована горизонтально, т. е. перпендикулярно солнечным лучам. В этом случае максимально улавливаются лучи, когда солнце находится в зените; 2) листовая пластинка ориентирована параллельно солнечным лучам, т. е. расположена более или менее вертикально. При этом растение лучше усваивает солнечные лучи в утреннее и предвечернее время; 3) листовые пластинки расположены по побегу диффузно, как у кукурузы, - то вертикально, то горизонтально. В этих случаях солнечная радиация улавливается наиболее полно в течение всего светового дня. Имеющиеся научные данные позволяют считать, что растения высоких широт, где преобладает низкое солнцестояние, чаще имеют вертикальную ориентацию листьев. При организации смешанных посевов, например кормовых трав, нужно обязательно учитывать структуру побегов компонентов этих посевов. Удачное сочетание кормовых трав с разной ориентацией листьев обеспечит больший урожай фитомассы.
Как уже отмечалось, в зависимости от недостатка или избытка освещенности многие растения способны размещать листья в плоскостях, перпендикулярной и параллельной направлению солнечных лучей, образуя так называемую листовую мозаику. Листовая мозаика образуется в результате рационального размещения неодинаковых по величине не только листовых пластинок, но и черешков. Типичную листовую мозаику можно наблюдать в фитоценозах с участием клена остролистного (Acer platanoides), липы сердцевидной (Tilia cordata), вяза обыкновенного (Ulmus laevis), ильма горного (Ulmus scabra) и других древесных пород. Четко прослеживается листовая мозаика у многих растений с горизонтальным размещением ветвей, например у плюща обыкновенного (Hedera helix) и многих травянистых растений (рис. 17).
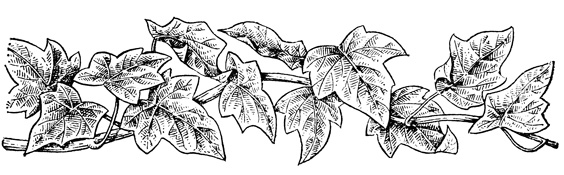
Рис. 17. Листовая мозаика у плюща (Hedera helix)
Компасные растения явно избегают сильного освещения. Их листовая пластинка расположена не перпендикулярно солнечным лучам, как у розеточных растений, а параллельно, как у эвкалиптов (Eucalyptus) или у дикого латука (Lactuca serriola), что предохраняет листья от перегрева в условиях избыточной солнечной радиации. Тем самым обеспечивается и благоприятное прохождение фотосинтеза и транспирации. Имеется целый ряд и других адаптивных приспособлений - как структурных, так и физиологических. Иногда же подобные приспособления носят явно сезонный характер. Такой пример хорошо иллюстрирует сныть обыкновенная (Aegopodium podagraria). В типичном местообитании, в дубравах, это растение в течение вегетационного периода производит два "поколения" листьев. Весной, когда почки деревьев еще не распустились и полог леса пропускает много света, сныть формирует листовую розетку. Эти листья по структуре (микро- и макроскопической) явно световые. Позднее, когда формируется густой полог леса и на поверхность почвы доходит лишь 3 - 4% лучистой энергии, появляется второе "поколение" листьев, явно теневых. Нередко можно наблюдать, что у одного отдельно взятого растения имеются одновременно световые и теневые листья. У шелковицы черной (Morus nigra) листья нижних ярусов кроны более крупные, лопастные, тогда как верхние ярусы кроны несут световые листья - более мелкие, лишенные лопастей. Лесообразующие породы подобным же образом формируют на периферии крону: в верхних ярусах - световые листья, а внутри кроны - теневые.
|
ПОИСК:
|
При использовании материалов проекта обязательна установка активной ссылки:
http://geoman.ru/ 'Физическая география'